An engram is a hypothetical spot in the brain where there is alleged to be a memory trace, an alteration in brain matter caused by the storage of memory. While scientists have claimed that there are countless engrams in your head, the notion of an engram has no robust scientific evidence behind it. No robust evidence for engrams has been found in any organism. Every study that has claimed to provide evidence for the existence of an engram has had problems that should cause us to doubt that good evidence for engrams was provided.
In a previous post I pointed out the not-really-science status of engrams by doing some queries on major preprint servers that store millions of scientific papers, servers such as the physics paper preprint server (which includes quanitative biology papers), the biology preprint server and the psychology preprint server. The queries (searching for papers that used the word "engram") showed only the faintest trace of scientific papers mentioning "engrams" in their titles. Only a handful of papers used that word in their title. An examination of such papers (discussed in my post) showed they provided nothing like any substantial evidence for the existence of any such thing as an engram.
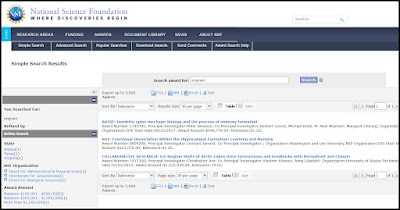
There is another way of testing whether this concept of engrams has any real observational support. We can use the grant search tool of the National Science Foundation. The National Science Foundation is a US institution that doles out billions of dollars each year in grants for scientific research. You can use the NSF's grant query tool to find out how much research money is being allocated to research particular topics.
You can perform the search by using the URL below:
https://www.nsf.gov/awardsearch/simpleSearchResult?queryText=engram
The results we get are the results shown below. We get only 3 matches. The last match is some climate paper having nothing to do with memory. So our search has produced only two National Science Foundation grants relating to the topic of engrams.
The second project ("Functional Dissociation Within the Hippocampal Formation: Learning and Memory") completed in 1992. Clicking on the link to the project, we see that $163,000 was spent, but no scientific papers are listed as resulting from the project.
The first grant is a grant of $996,778.00 (nearly one million dollars) that was given to a project entitled "Dendritic spine mechano-biology and the process of memory formation." The project started in 2017 and has a listed end date of July, 2022. The project description gives us a statement of speculative dogma regarding memory storage. There are actually very good reasons why the speculations cannot be correct. Below is the statement from the project description:
"The initiation of learning begins with changes at neuronal synapses that can strengthen (or weaken) the response of the synapse. This process is termed synaptic plasticity. Stimuli that produce learning lead to structural changes of the post-synaptic dendritic spine. The initial events of memory and learning include a temporary rise in calcium concentrations and activation of a protein called calmodulin. The next step is activation of calmodulin-dependent enzyme, kinase II (CaMKII). At the same time, structural rearrangements occur in the actin cytoskeleton leading to an enlargement of the spine compartment. How these initial events lead to remodeling of the actin cytoskeleton is largely unknown. This project focuses on the events that lead to the changes in actin cytoskeleton. The research also addresses the question of how these structural changes in the actin cytoskeleton are used to maintain memory."
To see why the main parts of the statement are not well-founded in observations, let us consider dendritic spines. A dendritic spine is a tiny protrusion from one of the dendrites of a neuron. The diagram below shows a neuron in the top half of the diagram. Some dendritic spines are shown in the bottom half of the visual. The bottom half of the visual is a closeup of the red-circled part in the top of the diagram.
An individual neuron in the brain may have about a thousand such dendritic spines. The total number of dendritic spines in the brain has been estimated at 100 trillion, which is about a thousand times greater than the number of neurons in the brain. The total number of synapses in the brain has also been estimated at 100 trillion. A large fraction of synapses are connected to dendritic spines.
Now, given such a high number of dendritic spines and synapses, we have the interesting situation that there is no possibility of correlating the learning of something and a strengthening of synapses or a strengthening or enlarging or growth of dendritic spines. Even if we are testing only a mouse, we still have an animal with trillions of dendritic spines and trillions of synapses. Scientists are absolutely unable to measure the size, strength or growth of all of those dendritic spines and synapses. The technology for doing that simply does not exist. What scientists can do is inspect a very small number of dendritic spines, taking snapshots of their physical state. But no such inspection would ever allow you to conclude that one or more dendritic spines had increased in size or grown or strengthened because some learning had occurred. Since dendritic spines slowly increase and decrease in size in an apparently random fashion, there is no way to tell whether the increase or decrease of a dendritic spine (or a small number of such spines) is being caused by learning or by the formation of a memory.
Therefore the statements below (quoted above) cannot be well-founded:
"The initiation of learning begins with changes at neuronal synapses that can strengthen (or weaken) the response of the synapse. This process is termed synaptic plasticity. Stimuli that produce learning lead to structural changes of the post-synaptic dendritic spine."
In fact, we know of the strongest reason why the hypothesis underlying such a claim cannot be true. The reason is that human memories are often extremely stable and long lasting, while dendritic spines and synapses are unstable, fluctuating things that have typical lifetimes of a few months or weeks. Read here to find some papers supporting such a claim. I can quote some scientists (Emilio Bizzi and Robert Ajemian) on this topic:
"If we believe that memories are made of patterns of synaptic connections sculpted by experience, and if we know, behaviorally, that motor memories last a lifetime, then how can we explain the fact that individual synaptic spines are constantly turning over and that aggregate synaptic strengths are constantly fluctuating? How can the memories outlast their putative constitutive components?"
The word "outlast" is a huge understatement here, for the fact is that human memories such as 50-year-old memories last very many times longer than the maximum lifetime of dendritic spines and synapses, and such memories last 1000 times longer than the protein molecules that make up such spines and synapses (which have average lifetimes of only a few weeks or less).
But enough of this long disputation of the claims made in the project description of the project entitled "Dendritic spine mechano-biology and the process of memory formation." Now let's look at what the million-dollar project has published so far in the way of results. We can see that by going to this page looking at the section entitled "PUBLICATIONS PRODUCED AS A RESULT OF THIS RESEARCH." The last three of these papers do not mention memory or engrams, so we may assume that they did nothing to substantiate claims about neural storage places of memory (engrams). The only paper mentioning memory or engrams in its title is a paper entitled "Exploring the F-actin/CPEB3 interaction and its possible role in the molecular mechanism of long-term memory." The paper can be read in full here.
The paper does not do anything to substantiate claims that memories are stored in engrams in the brain. The paper merely presents a speculative chemistry model and some speculative computer simulations. No experiments with animals have been done, and no research on human brains has been done. Apparently, there were no lab experiments of any type done, with all of the "experimentation" going on inside computers. The computer simulations do not involve the biochemical storage or preservation of any learned information. The quotes below help show the wildly speculative nature of the paper (I have put in boldface words indicating that speculation is occurring).
"Here we study the interaction between actin and CPEB3 and propose a molecular model for the complex structure of CPEB3 bound to an actin filament... Our model of the CPEB3/F-actin interaction suggests that F-actin potentially triggers the aggregation-prone structural transition of a short CPEB3 sequence....The CPEB/F-actin interaction could provide the mechanical force necessary to induce a structural transition of CPEB oligomers from a coiled-coil form into a beta-sheet–containing amyloid-like fiber...This beta-hairpin acts as a catalyst for forming intramolecular beta-sheets and could thereby help trigger the aggregation of CPEB3....These beta-sheets could, in turn, participate in further intermolecular interactions with free CPEB3 monomers, triggering a cascade of aggregation....Several possible mechanisms by which SUMOylation could regulate the CPEB3/F-actin interaction are discussed in SI Appendix....
We propose that SUMOylation of CPEB3 in its basal state might repress the CPEB3/F-actin interaction....Furthermore, the beta-hairpin form of the zipper suggests that it might be able to trigger extensive beta-sheet formation in the N-terminal prion domain, PRD....The beta hairpin form of zipper sequence is a potential core for the formation of intramolecular beta sheets... The maintenance of the actin cytoskeleton and synaptic strength then might involve the competition between CPEB3 and cofilin or other ABPs....We also propose that the CPEB3/F-actin interaction might be regulated by the SUMOylation of CPEB3, based on bioinformatic searches for potential SUMOylation sites as well as SUMO interacting motifs in CPEB3....We therefore propose that SUMOylation of CPEB3 is a potential inhibitor for the CPEB3/F-actin interaction."
The wildly speculative nature of the paper is shown by the boldface words above, and by the sentence at the end of the paper's long "Results" section: "Further experimental and theoretical work is required to determine which, if any, of these mechanisms is operating in neurons." Note well the phrase "which, if any" here. This is a confession that the authors are not sure a single one of the imagined effects actually occur in a brain.
In this case the US government paid a million dollars for essentially a big bucket of "mights" and "coulds," and the authors do not seem confident that any of these speculative effects actually occur in the brain. Whatever is going on here, it doesn't sound like science with a capital S (which I define as facts established by observations or experiments). Even if all of the wildly speculative "mights" and "coulds" were true, it still would not do a thing to show that memories lasting fifty years can be stored in dendritic spines and synapses that do not last for years, and are made up of proteins that have average lifetimes of only a few weeks. The idea that changes in synapse strength can store complex learned information has never made any sense. Information is physically stored not by some mere strengthening but by when some type of coding system is used to write information using tokens of representation. Never does a mere strengthening store information. The idea that you store memories by synapse strengthening makes no more sense than the idea that you learn school lessons by strengthening your arm muscles. If memories were stored as differences in synapse strengths, you could never recall such memories: because the brain lacks any such thing as a synapse strength reader.

No comments:
Post a Comment